
发布时间:2024-12-17 浏览:403次
短链脂肪酸(SCFAs)又称低级脂肪酸,是膳食纤维和抗性淀粉在肠道微生物发酵过程中产生的一类非常重要的天然无毒代谢产物。短链脂肪酸几乎在所有哺乳动物体内均有广泛分布,其种类包括甲酸、乙酸、丙酸、丁酸和戊酸等。主要产生 SCFAs 的菌群为厌氧菌,包括拟杆菌属、双歧杆菌属、梭菌属和链球菌属等 [1] 。
SCFAs 在维护肠道功能、维持葡萄糖稳态、改善食欲、调节能量代谢、提高免疫功能、抑制炎症以及抗肿瘤(尤其是结肠癌)等方面有积极的作用。SCFAs 不仅可以作为药物治疗疾病,甚至可以作为禽畜饲料以及化妆品的添加成分,具有提高动物抗氧化能力、增强免疫机能、提高生长性能和降低腹泻率,并可提高平均日增重及避免化妆品刺激成分导致的皮肤过敏反应 [2-6] 。本文总结了 SCFAs 维持生物体稳态的作用机制及其在禽畜生产中发挥的积极作用,为SCFAs的应用开发提供思路和参考。
1 短链脂肪酸维持生物体稳态的作用机制
SCFAs 不仅对胃肠道有保护作用,对肝脏、大脑、胰腺、心血管系统、皮肤组织和免疫系统等组织器官也产生积极的影响。
1.1 维持胃肠道稳态的作用机制
胃肠道细胞生理稳态是判断胃肠道受到外界侵袭的标志之一,因为健康胃肠道细胞以正常的方式生长、增殖、促进黏膜愈合并保持代谢稳态,能显著降低炎症形成、肿瘤侵袭和细胞迁移的机率。在胃肠道中,SCFAs 是调节能量代谢的关键因素,可激活 5'磷酸腺苷激酶(AMP)为代谢提供主要能量。

丁酸盐是肠上皮细胞的首选能源物质,实验证明人类结肠主要能量来自丁酸氧化而不是葡萄糖供能。丁酸盐还是最有效的细胞增殖抑制剂,是调节细胞增殖和分化的主要物质,可以调节 Treg/Th17 平衡,升高 IL-10、IL-12 和 TGF-并降低IL-17的水平。丁酸盐可在正常细胞中充当HAT激活剂,在癌细胞中充当 HDAC 抑制剂与 Foxo3 启动子,诱导结肠肿瘤细胞系凋亡,乙酸和丙酸盐还可增强丁酸盐在预防结肠癌方面的作用。丁酸可以使细胞周期阻滞在 G1 期,从而抑制细胞增殖。体内和体外实验都证明丁酸能够使相对氧化磷酸化水平升高、降低P-p27 和LC3-Ⅱ相对丰度、抑制结肠上皮细胞的自噬、加快结肠伤口愈合及维持人体结肠稳态 [7,8] 。口服丁酸还可抑制mTOR激酶活性和HDAC3 活性并促进巨噬细胞分化,以控制防御肽蛋白和 IgAmRNA 表达增加降低生物炎性反应 [9,10] 。
SCFAs 中乙酸和丙酸激活 GPR43 和丁酸激活GPR109a 被视为抑制胃肠道炎症的重要反应条件之一 [1] ,EHLJ7(一种四元黄连碱衍生物)能促进 SCFAs特别是丁酸的产生,进一步研究表明EHLJ7 可通过抑制 janus 激酶 2(JAK2)/信号转导和转录激活因子 3(STAT3)/细胞因子信号传导抑制子1 (SOCS1)通路的激活,降低TNF- 和IL-6 的水平与丁酸协同发挥抗溃疡性结肠炎的作用 [11] 。
1.2 维持肝脏稳态的作用机制
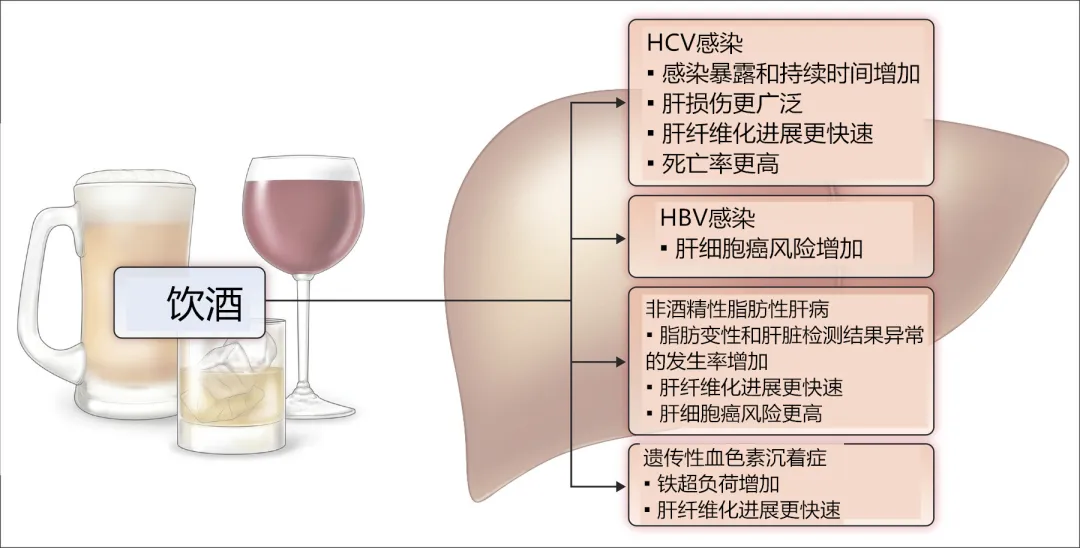
肝脏是机体重要的代谢器官,SCFAs 能够调节肝脏中的葡萄糖代谢。其中乙酸通过抑制 2,6-二磷酸果糖合成、激活糖异生和糖酵解,补充肝糖原起到保护肝脏的作用;丙酸可促进糖酵解,影响肝脏的葡萄糖代谢、抑制肝脏胆固醇的合成,保护肝脏 [1] 菊粉诱导小鼠肠道菌群产生的 SCFA(丙酸钠、丁酸钠和戊酸钠)可通过抑制 TLR-4/MyD88/NF- B 信号通路、抑制肝巨噬细胞M 1 和促进M 2 表达,导致TNF- 和iNOS水平降低、IL-10 和 Arg-1 水平上升来改善炎症,抑制酒精性肝病 [12] 。
1.3 维持大脑稳态的作用机制
肠可以通过肠源性激素的分泌将营养和能量状态的信号经脑肠轴传递到大脑皮层,而SCFAs可经肠系膜上静脉(SMV)和肠系膜下静脉(IMV)到达全身循环并且对一系列肠源性激素等起到调节作用。研究发现乙酸可以通过单羧酸转运蛋白穿过血脑屏障,这证明SCFAs可以作为信号分子将肠道状态传递给大脑 [1]
SCFAs 可以控制大脑小胶质细胞的功能并逆转小胶质细胞缺陷,有助于维持血脑屏障完整性并促进血脑屏障的发育。细菌疗法可逆转老年鼠脑中风模型体内短链脂肪酸含量下降,并逆转老年鼠大脑中的IL-17 和 T 细胞显著减少的现象,这说明 SCFAs 可减轻脑中风导致的的神经功能障碍和炎症,扭转老年小鼠脑中风恢复不良的状况 [1,13] 。朱慧越等 [14] 发现乙酸、丁酸和异丁酸能在不同程度上减轻小鼠抑郁样行为,其机制与SCFAs 修复下丘脑-垂体-肾上腺轴反馈调节障碍、恢复 5-羟色胺的正常代谢相关。
作为一种广谱抗癫痫药物的丙戊酸(VPA),因其价廉、患者耐受性强、副作用小,近几十年来一直被用于治疗癫痫以及双相情感障碍和偏头痛疾病。研究发现 VPA 可通过增强 -氨基丁酸的传递、减少 -羟丁酸的释放等方式发挥抗癫痫作用,并且VPA呈计量依赖性的增加 -H2AX,增加癌细胞对电离辐射的敏感性,阻止癌细胞 DNA 双链断裂修复,提高肿瘤细胞死亡率 [15] 。
1.4 维持胰腺稳态的作用机制
SCFAs可保护胰岛免受病毒侵害,能重塑LEW1.WR1大鼠的肠道微生物群,干扰Ⅰ型糖尿病(T1D)进程,降低因胰岛自身免疫障碍导致的 IFN- 、p40 和 STAT-1表达过量的现象 [16] 。乙酸可与 GPR43 结合增加骨骼肌细胞的自噬,是改善 II 型糖尿病(T2DM)介导的胰岛素抵抗(IR)过程中的关键调节因子,进而改善IR [17] 。
研究证明在小鼠饮食中补充不同比例的短链脂肪酸(乙酸钠、丙酸钠和丁酸钠)混合物干预 4 周后,可以显著降低小鼠胰岛素水平和空腹血糖水平(*P <0.05)[18] 其中丙酸钠可以显著降低血糖水平和胰岛素抵抗、增强胰岛 细胞功能、提高葡萄糖耐量,对胰腺组织损伤起到改善作用,而且灌胃丙酸钠可以改善高脂饮食引起的胰岛素抵抗(*P < 0.05),降低 IL-8 的水平,起到维持代谢健康和葡萄糖动态平衡的作用,对糖尿病起到缓解作用 [19]
1.5 维持心血管系统稳态的作用机制
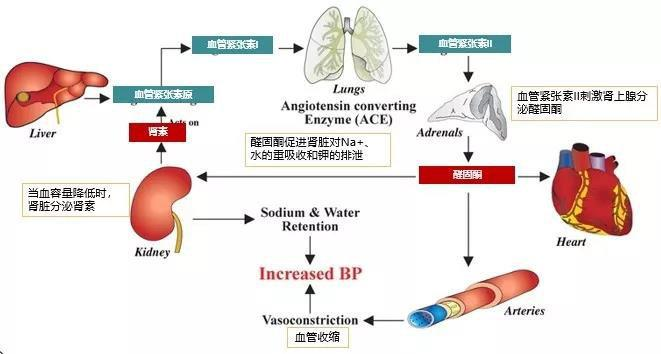
SCFAs可维持血压动态平衡,通过与Olfr78 (嗅觉受体 78)和 GPR41 (G 蛋白偶联受体 41)这两种短链脂肪酸受体结合,调节血管收缩和扩张实现对血压的调控 [20] 。丁酸盐可调节血管内皮细胞功能、降低其氧化应激程度和促炎因子的产生,改善动脉粥样硬化的症状 [21] 。丙戊酸通过刺激小鼠血管内皮细胞产生 t-PA(组织型纤溶酶原激活物),使小鼠体内血栓的形成几率下降 [22] 。异戊酸通过激活 PKA 介导的血管紧张素 a(ACh)诱发的纵向肌层平滑肌舒张 [23] 。短链脂肪酸(异丁酸、异戊酸)还能降低正常饮食雄性小鼠血清中三酰甘油水平,实现降血脂的效果 [24] 。
1.6 维持皮肤稳态的作用机制
皮肤很容易受到免疫系统的影响,许多炎症性皮肤病是由免疫介导的。细菌、古生菌、真菌、原生动物和病毒等共生体群落广泛分布在皮肤毛囊、皮脂腺和汗腺等位置,这些部位是皮肤免疫中免疫细胞和共生体相互作用的关键部位,这些共生微生物不仅控制肠道炎症,还控制皮肤、口腔、呼吸道和泌尿生殖道等外部屏障器官。

SCFAs是皮肤Tregs的有效调节剂,皮肤共生体会产生 SCFAs。研究发现皮下注射或局部应用丁酸钠(SB)可使 Treg 特异性转录因子 Foxp3 和 IL-10的编码基因上调,显著降低半抗原致敏小鼠的过敏反应,因此可以推测,短链脂肪酸可用于治疗皮肤炎症反应 [6] 。
1.7 维持免疫系统稳态的作用机制
短链脂肪酸能够选择性地影响肌肉、肝脏和脂肪组织中的葡萄糖储存和能量代谢。SCFAs 能够诱导抗菌肽的产生并调节调节性 T 细胞(Tregs)的数量和功能使得 Foxp3+、IL-17A、IL-1、IL-6、IL-12 和 IL-18 表达增加,表明 SCFAs 有助于调节宿主免疫应答。
乙酸盐灌胃肥胖和糖尿病大鼠可改善其葡萄糖耐受量和体重增加的现象,并且 HFD 小鼠喂食富含丁酸盐的饲料会增加小鼠产热和能量消耗,并抵抗肥胖。乙酸通过改变脂质代谢并减少淋巴组织中 T 细胞影响大脑中枢系统并降低宿主食欲,表现出对糖尿病患者的高度保护作用 [25] 。然而过量应用 SCFAs 可能会干扰离子依赖性转运体的正常表达,例如过量应用乙酸和丙酸会使细胞内弱酸的离解增强导致H + 浓度上升,从而影响Na-H、Na-K、H-K、Na-Ca 交换、Na/K-atp 酶 H/K-atp 酶活性对生物的生理稳态平衡起到负作用 [26] 。
1.8 维持肾脏稳态的作用机制
SCFAs 与肾脏之间有着的紧密联系,外源性补充SCFAs 能延缓慢性肾脏病患者(CKD)发生和发展。SCFAs 通过与 G 蛋白偶联受体结合,抑制组蛋白去乙酰化酶调节肾素血管紧张素系统和细胞自噬、抑制 NF- B 通路和MAPK 通路,促进TNF- 、IL-1、IL-6和iNOS水平下调等方式起到延缓肾脏炎症和纤维化的作用,使得SCFAs 成为肾脏疾病治疗的新研究方向 [27] 。
1.9 维持支气管系统稳态的作用机制
SCFAs 与预防食物过敏和哮喘有关,其可激活中性粒细胞和巨噬细胞上的 FFA2 和 FFA3 受体,减少气道炎症期间气道中 IL-8 的产生与治疗所需糖皮质激素的使用量,并通过激活ERK1/2 和FOXP3 的表达增加血清中 IL-4、IL-10 的含量、降低血清中 TGF- 1和 IL-2 的水平,减轻肺部过敏性患者的炎症反应。动物实验发现,小鼠口服乙酸、丙酸或丙酸酯、丁酸能显著降低小鼠过敏性气道炎症的严重程度 [1, 28] 。
2 在禽畜生产中的应用
在饲料中添加 SCFAs 可显著提高仔猪血清中甘油三酯和磷的含量,并增加仔猪日增重(ADG)和平均日采食量(ADFI)[2] 。SCFAs不但可以降低仔猪肠道中pH、减少细胞凋亡,还可以增加仔猪肠道中黏蛋白MUC1、MUC2、IGF-1、IGF-IR、GLP-2 和GLP-2R的相对mRNA 表达,并降低Bax、Caspase-3 表达和血清中促炎细胞因子TNF- 、IL-6、IL-1、IL-8 和IL-1 的水平降低仔猪腹泻率。
丁酸钠还降低血清中丙二醛含量,提高 IFN- 含量,从而提高断奶仔猪与幼鸡的机体抗氧化能力和增强免疫机能,提高生长性能、降低腹泻率并提高平均日增重 [3,4] 。Liu 等 [5] 研究发现口服丁酸钠可以下调 IG-FBP-3、Caspase-3 和Bcl-2 等mRNA的表达、上调IG-FBP-5、IGF-1R 和CDK6 等mRNA 的表达,改善断奶羔羊的动物生长性能。
3 总结与展望
深入了解 SCFAs 的作用可为维持生物体稳态提供有用信息,SCFAs 是肠道微生物区系产生的,对机体有正向调节作用的一类脂肪酸,在抑制疾病产生、发展中起到重要作用。本文讨论了短链脂肪酸维持生物体生理状态稳定及禽畜生产的保护作用。
全民大健康时代益生菌代谢产生的短链脂肪酸成为关注热点,了解 SCFAs 的功能作用,有望为影响宿主健康的微生物代谢物作用提供一般规律。但从目前的研究来看单用 SCFAs 治疗疾病的疗效不是十分显著,而且关于SCFAs调控细胞自噬的具体调控机制尚不完全清楚。应考虑使用SCFAs与抗炎药、益生菌(如菊粉)等结合来治疗疾病,希望本文会为 SCFAs 的应用提供一个新的思路。
#参考文献#
[ 1 ] HU J, LIN S, ZHENG B, et al.Short-chain fatty acids in control of energy metabolism [ J ] . Crit Rev Food Sci Nutr, 2018, 5 ( 8 ) :1243-1249.
[ 2 ] 寇莎莎,王诏升,徐德旺,等. 日粮中添加不同水平丁酸钠对断奶仔猪生长性能、腹泻率及血液生化指标的影响 [ J ] . 中国畜牧兽医,2018,45(7):1841-1848.
[ 3 ] DIAO H, JIAO A R, YU B, et al. Gastric infusion of short -cha-in fatty acids can improve intestinal barrier function in weaned piglets [ J ] . Genes Nutr, 2019, 14 ( 1 ) : 4.
[ 4 ] 赵瑜,靳文广,STEPHAN BAUWENS. 丁酸盐对幼龄鸡的功效 [ J ] . 国外畜牧学(猪与禽),2016,36(2):52-54.
[ 5 ] LIU L, SUN D, MAO S, et al. Infusion of sodium butyrate pro-motes rumen papillae growth and enhances expression of ge-nes related to rumen epithelial VFA uptake and metabolism in neonatal twin lambs [ J ] . Journal of Animal Science, 2019, 97( 2 ) : 909-921.
[ 6] SCHWARZA,BRUHS A, SCHWARZT.TheShort-Chainfatty acid sodium butyrate functions as a regulator of the skin im-mune system [ J ] . J Invest Dermatol, 2017, 137 ( 4 ) : 855-864.
[ 7 ] ZENG H, TAUSSIG DP, CHENG WH, et al. Butyrate inhibits cancerous HCT116 colon cell proliferation but to a lesser ex-tent in noncancerous NCM460 colon cells [ J ] . Nutrients, 2017, 9( 1 ) : 25.
[ 8 ] KAIKO GE, RYUSH,KOUES OI, et al. The colonic crypt pro-tects stem cells from microbiota-derived metabolites [ J ] . Cell,2016, 165 ( 7 ) : 1708-1720.
[ 9 ] JIRSOVA Z, HECZKOVA M, DANKOVA H, et al. The Effect of butyrate-supplemented parenteral nutrition on intestinal de-fence mechanisms and the parenteral nutrition-induced shift inthe gutmicrobiota in the rat model [ J ] . Bio Med Research Inter-national, 2019, 14 ( 1 ) : 7084734.
[ 10 ] SCHULTHESS J, PANDEY S, CAPITANI M, et al. The short chain fatty acid butyrate imprints an antimicrobial program in macrophages [ J ] . Immunity, 2019, 50 ( 2 ) : 432-445.
[ 11 ] TANG X, LI X, WANG Y, et al. Butyric acid increases the therapeutic effect of EHLJ7 on ulcerative colitis by inhibiting JAK2/ STAT3/SOCS1 signaling pathway [ J ] . Front Pharma-col, 2019, 10: 1553.
[ 12 ] 王振. 菊粉经短链脂肪酸调节巨噬细胞极化对酒精性肝病的抗炎作用研究 [ D ] . 银川:宁夏医科大学,2020.
[ 13 ] LEE J, D’AIGLE J, ATADGA L, et al. Gut microbiota-deriv-ed short-chain fatty acids promote post-stroke recovery in aged mice [ J ] . Circ Res, 2020, 127 ( 4 ) : 453-465.
[ 14 ] 朱慧越,邹仁英,许梦舒,等.短链脂肪酸-酰化淀粉对小鼠抑郁样行为的缓解及机制 [ J ] . 食品与发酵工业,2021,47(6):26-33.
[ 15 ] YARMOHAMADI A, ASADI J, GHARAEI R, et al. Valproicacid,a histone deacetylase inhibitor,enhances radiosensitivity in breast cancer cell line [ J ] . Cancer Res, 2018, 9 ( 2 ) : 86-92.
[ 16 ] NEEDELL JC, IR D, ROBERTSON CE, et al. Maternal treat-ment with short-chain fatty acids modulates the intestinal microbiota and immunity and ameliorates type 1 diabetes in the offspring [ J ] . PloS One, 2017, 12 ( 9 ) : e0183786.
[ 17 ] LINGY, HAIQI L, WENTAO LIN,et al.Exercise ameliorates insulin resistance of type 2 diabetesthrough motivating Short-Chain fatty Acid-Mediated skeletal muscle cell autophagy [ J ] .Biology, 2020, 9 ( 8 ) :203.
[ 18 ] 刘倩,陈成,辛鑫,等.短链脂肪酸对高脂饮食诱导肥胖小鼠糖脂代谢紊乱的影响 [ J ] . 肝脏,2018,23(7):591-595.
[ 19 ] CHAMBERSES,BYRNECS,MORRISONDJ, et al.Dietarysupplementation with inulin-propionate ester or inulin im-proves insulin sensitivity in adults with overweight and obes-ity with distinct effects on the gut microbiota, plasma meta-bolome and systemic inflammatory responses: a randomized cross-over trial [ J ] . Gut, 2019, 68 ( 8 ) : 1430-1438.
[ 20 ] MIYAMOTO J, KASUBUCHI M, NAKAJIMA A, et al. The role of short-chain fatty acid on blood pressure regulation [ J ] .Curr Opin Nephrol Hypertens, 2016, 25 ( 5 ) : 379-383.
[ 21 ] HOFFMAN JB, PETRIELLO MC, HENNIG B. Butyrate modulates Cav-1 and its binding partner AhR, leading to dif-ferential Cyp1a1 and Cyp1b1 gene expression in vascular en-dothelial cells [ J ] . Faseb J, 2016, 30 ( 1 ) . 1175.
[ 22 ] LARSSON P, ALWIS I, NIEGO B, et al. Valproic acid selec-tively increases vascular endothelial tissue-type plasminogen activator production and reduces thrombus formation in the
mouse [ J ] . J Thromb Haemost, 2016, 14 ( 12 ) : 2496-2508.
[ 23 ] BLAKENEY BA, CROWE MS, MAHAVADI S, et al. Bran-ched short-chain fatty acid isovaleric acid causes colonic smooth muscle relaxation via cAMP/PKA pathway [ J ] . Dig Dis Sci, 2019, 64 ( 5 ) : 1171-1181.
[ 24 ] 谭文倩,李锐,于鑫,等.低剂量黄连素对小鼠肠道短链脂肪酸和胆汁酸成分以及血脂的影响 [ J ] . 复旦学报(医学版),2020,47(1):93-100.
[ 25 ] MARINO E, RICHARDS JL, MCLEOD KH, et al. Gutmicrobial metabolites limit the frequency of autoimmune Tcells and protect against type 1 diabetes [ J ] . Nat. Immunol,2017, 18 ( 8 ) : 552-562.
[ 26 ] HE X, LU Z, MA B, et al. Chronic heat stress damages small intestinal epithelium cells associated with the adenosine 5'- Monophosphate-Activated protein kinase pathway in broilers [ J ] . J. Agric. Food Chem, 2018, 66 ( 28 ) . 7301-7309.
[ 27 ] 罗科娜,蔡珂丹,罗群. 肠道菌群代谢产物短链脂肪酸在慢性肾脏病中的研究进展 [ J ] .中国微生态学杂志,2020,32 (8):983-987.
[ 28 ] HALNES I, BAINES KJ, BERTHON BS, et al. Soluble fibre meal challenge reduces airway inflammation and expression of GPR43 and GPR41 in asthma [ J ] . Nutrients, 2017, 9 ( 1 ) : 57.
#酒局守护神 #酉神1号 #酉神一号 #酉神壹号 #酒神1号 #酒神一号 #酒神壹号 #酉神1号解酒液 #酉神1号有机酸解酒液



